
Travis Christofferson, Tripping Over The Truth (2019).
Energy production in normal cells takes place primarily in cellular organelles called mitochondria. Mitochondria generate energy in the form of ATP (adenosine triphosphate) through a highly efficient process called oxidative phosphorylation, which is dependent on the presence of oxygen. Cancer cells have a markedly reduced capacity to generate energy via this mitochondrial pathway and an increased ability for energy production via the less efficient pathway of glycolysis (glucose fermentation). The marked increase in glycolysis in cancer cells is mediated by a massive overproduction of an embryonic form (HK2) of hexokinase, a key enzyme driving this energy pathway. Cancer cells also produce energy through mitochondrial substrate-level phosphorylation (mSLP), a glutamine pathway that can produce ATP in a much less efficient manner than oxidative phosphorylation in healthy cells. The dependency on glucose fermentation and glutamine metabolism is a critical weakness of all cancer types and a potential Achilles heel that provides opportunity for a highly focused but broadly applicable approach to treatment.
The striking difference in energy production in normal vs. cancer cells has a morphologic (structural) equivalent. Mitochondria of healthy cells have abundant cristae seen as parallel tubular structures within the mitochondrion (left figure below) which are markedly decreased in cancer-cell mitochondria (right figure below).

High-resolution image of a healthy mitochondrion. Note the tubular structures (cristae) inside the mitochondrion. The cristae are where energy is produced via oxidative phosphorylation in normal cells.
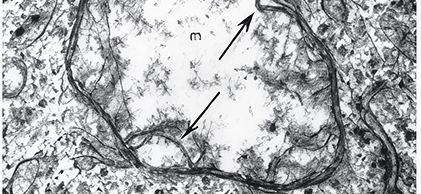
High-resolution image of a mitochondrion from a cancer cell. Note the almost complete absence of cristae (arrows) and rarefied matrix (m), features consistent with mitochondrial injury and diminished oxidative phosphorylation.
The dependence of cancer cells primarily on glucose fermentation for ATP production has been exploited in the development of current tumor imaging technologies that use labeled glucose analogs. Specifically, Positron Emission Tomography (PET) scans identify the distribution and relative size of primary and/or metastatic sites of a tumor within the body of the patient. These scans are critical to not only cancer detection but its management. In the image from a PET scan to the right, there is stunning visual evidence of the voracious appetite tumor cells have for glucose compared to normal tissue. The reddish variably sized and shaped sites within the scan represent metastasis of a tumor and are visible because of the disproportional uptake of the glucose analog by cancer cells relative to the surrounding normal tissue.
The dependence of cancer cells primarily on glucose fermentation for ATP production has been exploited in the development of current tumor imaging technologies that use labeled glucose analogs. Specifically, Positron Emission Tomography (PET) scans identify the distribution and relative size of primary and/or metastatic sites of a tumor within the body of the patient. These scans are critical to not only cancer detection but its management. In the image from a PET scan below, there is stunning visual evidence of the voracious appetite tumor cells have for glucose compared to normal tissue. The reddish variably sized and shaped sites within the scan represent metastasis of a tumor and are visible because of the disproportional uptake of the glucose analog by cancer cells relative to the surrounding normal tissue.
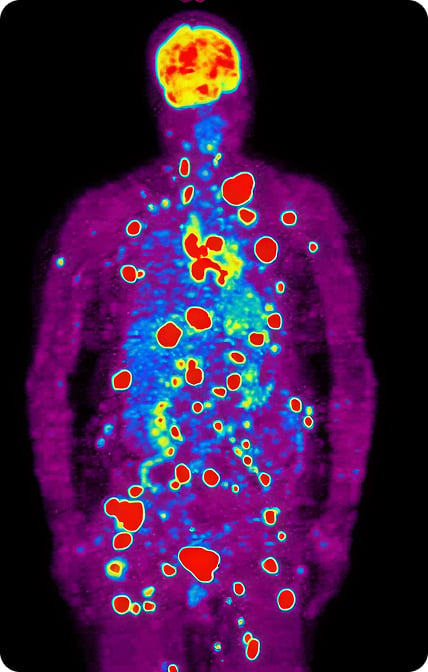
Positron Emission Tomography (PET) scan.
The relationship between mitochondrial dysfunction and genetic instability is very complex and likely bi-directional. However, there is growing body of peer-reviewed scientific evidence that supports once a cell has an impaired ability to produce energy through oxidative pathways, the genomic instability (increased potential for DNA mutations to occur) that accompanies tumor development, inevitably follows. Many initiating agents in cancer including chemicals, contact irritants, viruses, radiation, inflammation, etc. can directly cause mutations to DNA. It is also true these and other reactive or cytotoxic compounds can damage mitochondria directly. These agents can also cause altered mitochondrial function through mutations of mitochondrial and/or nuclear genes regulating mitochondrial structure and function. The metabolic theory of cancer states that once the mitochondria of a given cell become dysregulated, and the cell reverts to glucose fermentation to obtain energy, a plethora of metabolic and epigenetic modifications can occur and predispose to the development of cancer.
This metabolic theory of cancer has evolved over time into a model that integrates the stem cell theory into the metabolic theory. This integrated theory suggests that cancer originates from chronic insufficiency of oxidative phosphorylation in stem cells leading to the formation of cancer stem cells with abnormal energy metabolism, which ultimately leads to malignancy. Within the cancer stem cell, respiratory impairment (reduced oxidative phosphorylation) not only shifts these cells to glucose fermentation and glutamine energy metabolism but also induces overexpression of oncogenes and inactivation of tumor suppressor genes, which contribute to abnormal energy metabolism and neoplastic transformation.
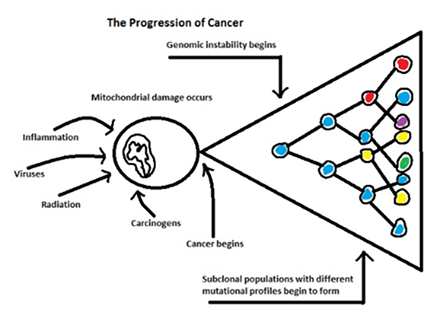
Metabolic Theory of Cancer Progression
The diagram above depicts, in an overly simplified manner, the metabolic theory of cancer progression. An inciting agent initiates the process which results in direct mitochondrial toxicity and/or alterations in gene expression involving mitochondrial structure and function. Once a threshold of mitochondrial dysregulation and concomitant epigenetic changes is reached, uncontrolled proliferation begins. The different colors in the figure above depict cells with different mutational profiles within the same tumor. If an oncologic drug targets a specific mutation, those subclonal populations of cancer cells with non-targeted mutations will be resistant to treatment.
The mitochondrial dysregulation/dysfunction and primary dependence on enhanced glucose fermentation for ATP production in cancer cells provide opportunity for multi-faceted treatment protocols utilizing dietary intervention, repurposed (off-label) drugs, vitamins, and other supplements that exploit cancer-cell dependence on glucose and glutamine for energy production. This metabolic approach to cancer treatment can serve as an alternative or adjunct therapy to conventional chemo-, radio-, and hormonal cancer therapies.
Metabolically directed therapies markedly diminish injury to normal cells in contrast to conventional and targeted chemotherapies and immunotherapies which are generally cytotoxic to both cancerous and normal cells in susceptible tissues. Furthermore, conventional cytotoxic chemotherapy is known to create a microenvironment of hypoxia and inflammation within the treated tumor(s), conditions that favor local spread and metastasis of resistant populations of cancer cells and cancer stem cells. Cancer stem cells, unlike rapidly proliferating cancer cells, are resistant to all conventional chemotherapeutics and play a major role in cancer recurrence. So even though a patient’s cancer is in remission and undetectable by imaging, there is a significant potential for recurrence from chemotherapeutic-resistant cancer stem cells.
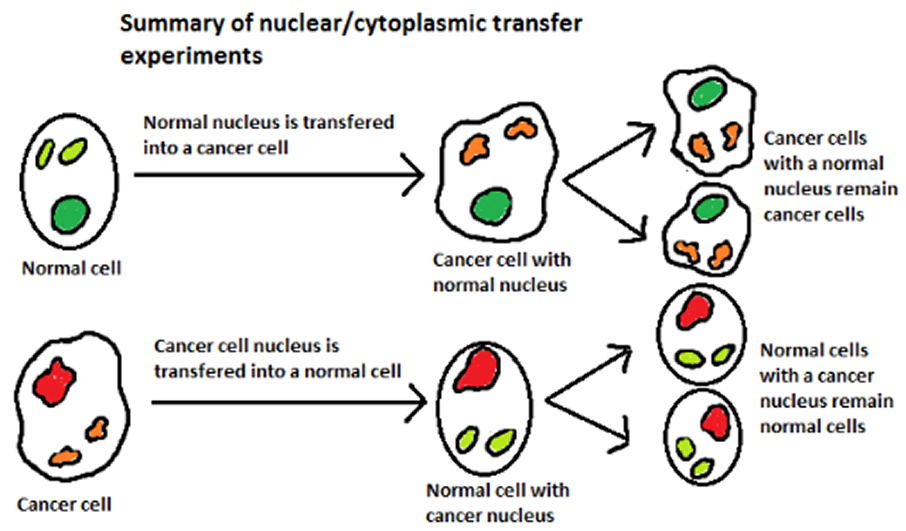
A series of simple yet elegant nuclear transfer experiments preformed in the mid 1980’s provided early evidence that metabolic and epigenetic factors are important drivers of cancer. When nuclei of cancer cells were transferred into normal cells, the cells remained normal. However, when nuclei of normal cells were transferred into cancer cells, surprisingly, the cells maintained the malignant phenotype. These simple experiments illustrate the potential for cytoplasmic and/or specific mitochondrial alterations rather than mutations to nuclear DNA as critical to transformation of normal cells to cancer cells.
Experimental evidence supports that both rapidly dividing cancer cells and cancer stem cells are susceptible to several repurposed drugs including ivermectin and the benzimidazoles whose anti-cancer mechanisms, in large part, involve inhibition of glucose fermentation and glutamine metabolism. There is clear evidence of efficacy and excellent tolerability in multiple case reports of cancer patients treated with these repurposed drugs, often in combination with a ketogenic diet. Most of these cases involve intervention in late-stage cancers resistant to conventional oncologic therapy. Remarkable results, including apparent cures, with no or little evidence of toxicity have been described. Much of the mainstream oncology community is dubious of the clinical evidence given its anecdotal nature and the absence of data from conventional well-designed clinical trials supporting efficacy and safety of these drugs in cancer treatment. This presents a conundrum to advancing these off-patent repurposed drugs as cancer treatments through an FDA approval process since no pharmaceutical company will incur the huge expense of clinical development of a drug that’s non-patentable. Nevertheless, a growing number of physicians, including oncologists, worldwide are exploring the off-label use of these drugs and others in combination with dietary intervention and relevant vitamins and supplements for the treatment of early to late-stage cancer malignancies.